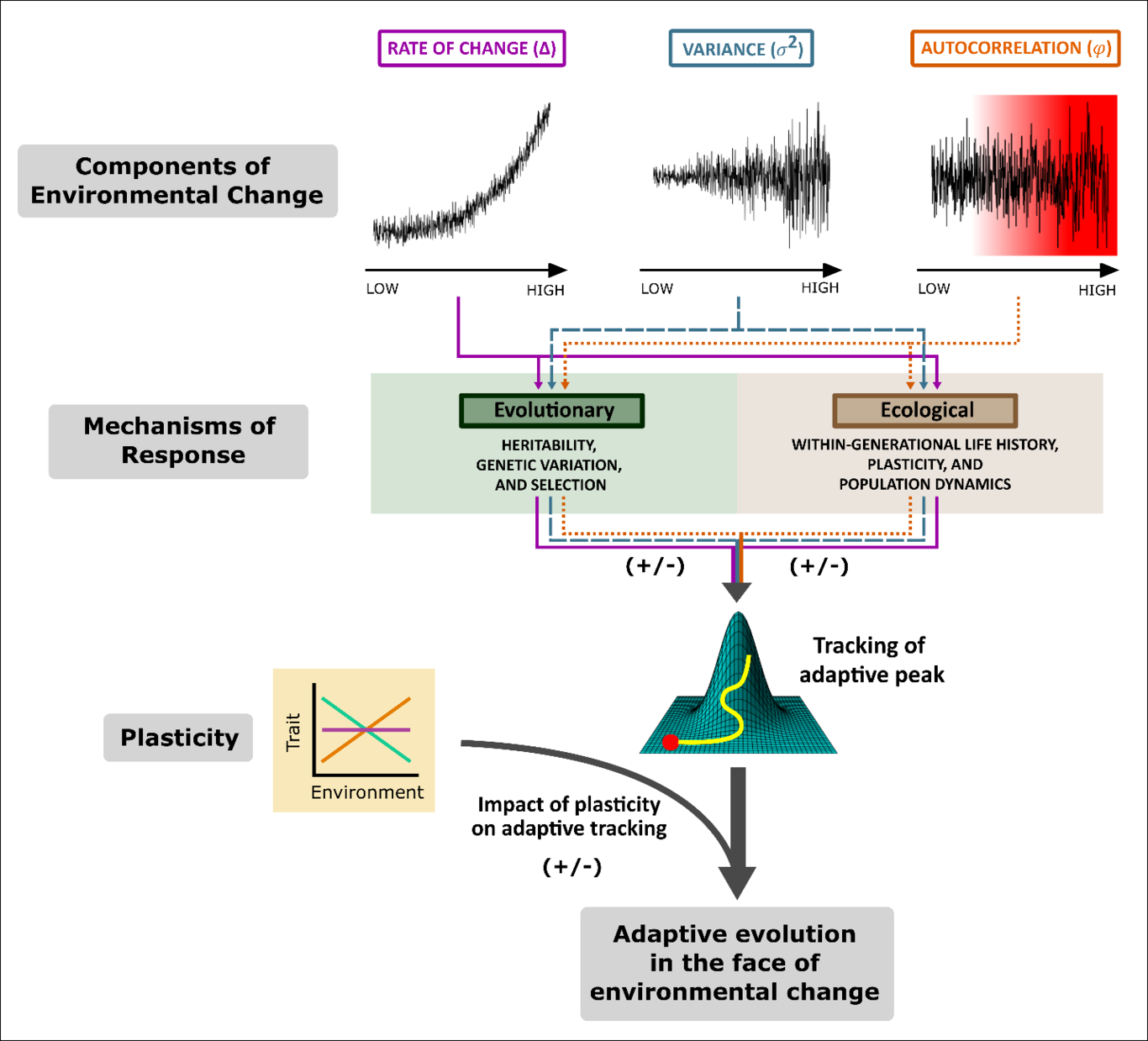
1 IntroductionUnderstanding, quantifying, and predicting the ability of organisms to adapt to changing environments is at the core of eco-evolutionary research[1,2]. In the face of unprecedented environmental change, natural populations, especially those with limited mobility, can avoid extinction via phenotypic plasticity and/or adaptive evolution [3]. However, our understanding of the interplay between selection and plasticity in changing environments is surprisingly limited[4–8]. This limitation is not trivial, for plasticity can itself evolve[9], can be adaptive or nonadaptive[10], and has seemingly contradictory effects on adaptive evolution[11], on which we focus here. For decades, researchers have theorized whether plasticity facilitates or hinders adaptive evolution[9,12]; the evidence is contradictory and general patterns have not emerged [5,10,11,13,14].The primary conflicting hypotheses for whether plasticity facilitates or hinders adaptive evolution are:(H1) plasticity weakens directional selection by masking genotypic variation (Bogert Effect [15]), thus slowing the rate of genetic change[5,16–18] vs.(H2) plasticity facilitates evolution by allowing the population to persist under environmental change long enough for genetic change to occur[19–22] (Plasticity-First Hypothesis [21] orBaldwin Effect [19]).This debate remains unresolved, for even when theoretical predictions agree with empirical findings[5,10,11,13,14,23], we lack a general framework to ascertain the context-dependency of the prevalent mechanism. Here, we introduce a framework based on environmental change context, to outline clear null hypotheses for when and how plasticity interacts with directional evolution. We place the plasticity facilitates vs. hinders selection debate on two ends of a continuum, and specify the properties of environmental change–rate of mean change , variability , andtemporal autocorrelation –that influence how plasticity impacts adaptive evolution.The type of environmental change a population experiences can alter its likelihood of adaptation and, ultimately, persistence[24–27]. Studies of demographic[28], genetic[29], and evolutionary rescue[30], show that rate of mean change, variability, and temporal autocorrelation of a population’s selective environment impact population persistence[24,25,29,31–35]. However, because different types of environmental change can have contradictory effects on plasticity and evolution[34,36–38], elucidating these dynamics is not trivial. Consequently, there is an urgent need to place this discussion on the environmental stage in a generalizable way that will allow ecologists and evolutionary biologists to better contextualize, mechanistically understand, predict, and compare their findings.Moving optimum theory links environmental change to the resulting evolutionary responses. Three decades of research on this theory shows that, when a population is confronted with an environment that changes directionally, there is a critical rate of changethat must be matched by change in the mean phenotype of the population, such that the mean remains close to the theoretical phenotypic optimum . In this context, a phenotypic lag between the mean phenotype and the optimum phenotype typically emerges which, if too large, makes extinction certain [39–41]. Evolutionary (e.g. , selection, genetic variation) and ecological processes (e.g. , within-generation life history, plasticity and population dynamics) together influence the limit of how far a population can lag without going extinct. The contribution of plasticity to population persistence and adaptation is largely determined by this phenotypic lag: how much of the short- or long-term lag can be compensated for or even hindered by plasticity?We argue that hypotheses such as the Bogert Effect and the Plasticity-First Hypothesis / Baldwin Effect are not mutually exclusive. Rather, plasticity may facilitate or hinder adaptive evolution depending on the properties of environmental change. To assess the impact of plasticity on the ability of a population to evolutionarily track a changing environmental optimum, we specify the links among the type of environmental change, plasticity, and adaptive evolution by considering several fundamental processes. Thus, we utilize both theoretical and experimental studies to:Assess how three key components of environmental change (rate of mean change , variability , and temporal autocorrelation ) each alter the mechanisms behind phenotypic tracking of a moving optimum ([i] Genetic variation, heritability, and selection , and [ii] life history, plasticity andpopulation dynamics ).Introduce a unified framework of testable hypotheses detailing how those three components of environmental change can influence the relative benefit of plasticity to adaptive evolution.